

Retatrutide Structural Mechanism: What Cryo-EM Reveals About Triple-Receptor Agonism
A single peptide that fits three different receptor locks simultaneously, that is the central engineering feat behind retatrutide. Understanding the Retatrutide Structural Mechanism: What Cryo-EM Reveals About Triple-Receptor Agonism requires stepping inside the molecular architecture of a 39-amino acid chain and asking a precise question: how does one molecule activate the GLP-1 receptor, the GIP receptor, and the glucagon receptor at the same time without losing potency at any of them? Cryo-electron microscopy (cryo-EM) has now provided detailed answers, and those answers explain why retatrutide behaves so differently from earlier incretin-based therapies.
Key Takeaways
- Retatrutide adopts a single continuous alpha-helix conformation when binding to all three target receptors, a structural uniformity confirmed by cryo-EM.
- Non-canonical amino acids at specific positions protect the peptide from enzymatic degradation and fine-tune receptor selectivity.
- The N-terminal segment drives receptor activation by penetrating the transmembrane core, while the C-terminal segment governs selectivity through extracellular interactions.
- Retatrutide is roughly 8.9 times more potent at the GIP receptor than native GIP, while its glucagon receptor activity is intentionally moderated to limit hyperglycemia risk.
- A fatty acid side chain enables albumin binding, extending the half-life to approximately six days and supporting once-weekly dosing.
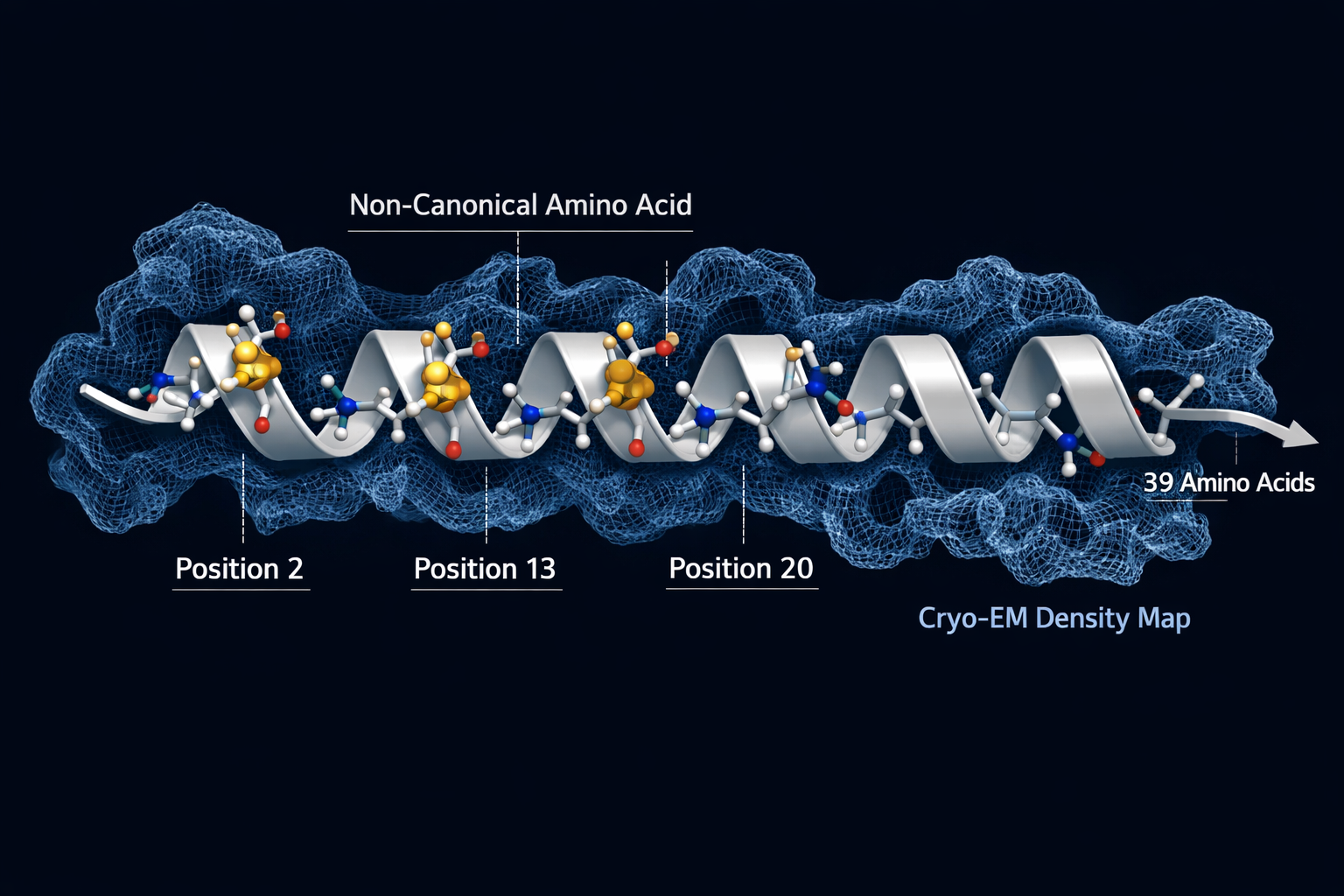
The Alpha-Helix Architecture Behind Triple-Receptor Binding
The most striking finding from cryo-EM studies is structural simplicity at the core. Despite engaging three pharmacologically distinct receptors, GLP-1R, GIPR, and GCGR, retatrutide maintains a single continuous alpha-helix conformation across all three binding events. This is not a trivial achievement. Most peptide ligands adopt slightly different conformations depending on the receptor environment they encounter. Retatrutide's rigid helical backbone allows it to slot into each receptor's binding pocket without requiring a structural reset.
This conformational consistency is not accidental. The peptide's sequence was engineered to include non-canonical amino acids that lock the helix in place:
- Alpha-aminoisobutyric acid (Aib) at positions 2 and 20, resists degradation by dipeptidyl peptidase-4 (DPP-4), the enzyme that rapidly breaks down native GLP-1.
- Alpha-methyl-L-leucine at position 13, supports GIP receptor activity and contributes to helical stability.
These modifications are part of what separates retatrutide from earlier GLP-1 peptide generations that lacked this level of structural engineering.
"The rigid alpha-helical backbone of retatrutide is not a byproduct of its design, it is the design."
The peptide also carries a fatty acid side chain that binds albumin in circulation, extending its half-life to roughly six days. This pharmacokinetic feature, combined with its enzymatic resistance, supports a once-weekly dosing schedule, a significant practical advantage over shorter-acting compounds.
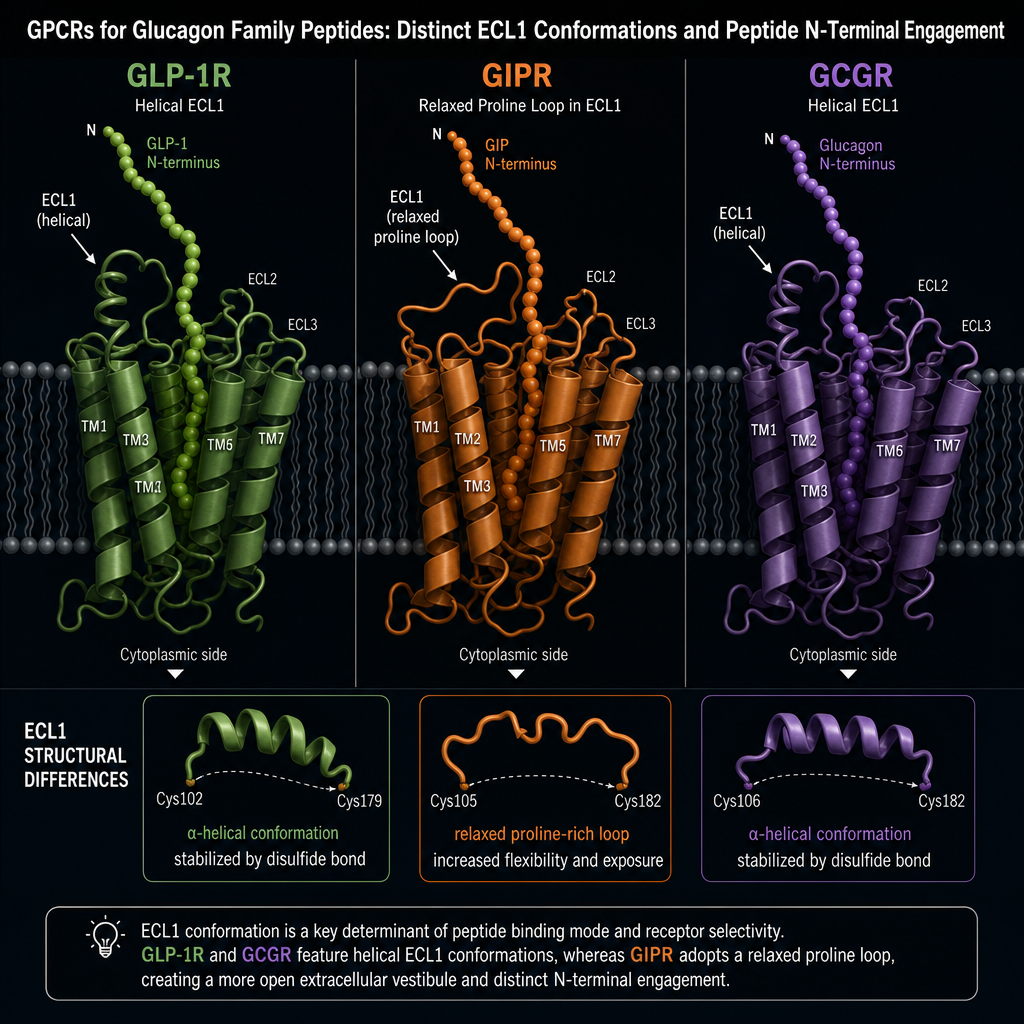
How Cryo-EM Maps the Retatrutide Structural Mechanism Across Three Receptors
Cryo-EM resolved the bound structures of retatrutide at each of its three target receptors, revealing a consistent two-part binding strategy:
| Segment | Residues | Primary Interaction |
|---|---|---|
| N-terminal | 1 to 13 | Penetrates transmembrane domain core |
| C-terminal | 14 to 30 | Engages extracellular regions |
The N-terminal segment is the activation trigger. It inserts into the hydrophobic core of each receptor's transmembrane bundle, initiating the conformational change that signals downstream G-protein coupling. The C-terminal segment is the selectivity filter, making contact with extracellular loops that differ between receptor subtypes.
One notable receptor-specific difference involves extracellular loop 1 (ECL1). In GLP-1R and GCGR, ECL1 adopts a helical structure. In GIPR, ECL1 takes a relaxed loop conformation because of proline residues in that region. Retatrutide accommodates this difference without altering its core helical shape, a testament to the design flexibility built into its sequence.
For researchers exploring dual receptor agonism mechanisms, this structural data illustrates precisely why adding a third receptor target requires more than simply extending a peptide chain.
Potency Profile and Metabolic Consequences of Triple-Receptor Agonism
Understanding the Retatrutide Structural Mechanism: What Cryo-EM Reveals About Triple-Receptor Agonism is incomplete without examining what each receptor activation actually does metabolically:
- GLP-1R activation, suppresses appetite and slows gastric emptying, reducing caloric intake.
- GIPR activation, enhances glucose-dependent insulin secretion and influences adipose tissue metabolism.
- GCGR activation, increases energy expenditure through hepatic lipid oxidation and thermogenesis.
Retatrutide's potency is deliberately asymmetric. It is approximately 8.9 times more potent at GIPR than native GIP, amplifying the insulin-sensitizing and fat-mobilizing effects of that receptor. At GCGR and GLP-1R, it operates at roughly 0.3 to 0.4 times the potency of endogenous glucagon and GLP-1, respectively. This deliberate moderation at GCGR limits the hyperglycemia risk that full glucagon activation would otherwise carry.
This potency calibration helps explain why clinical data show retatrutide producing 4 to 8 percent more weight loss than dual GLP-1/GIP agonists at comparable doses. The added glucagon receptor contribution raises resting energy expenditure in ways that appetite suppression alone cannot achieve.
Researchers interested in how incretin-based peptides compare across generations can explore GLP-1 incretin research themes for broader context. Those examining metabolic peptide research may also find value in reviewing body composition research themes related to tesa, which targets a different but metabolically relevant pathway. For a direct look at the compound itself, the GLP-3 retatrutide research product page provides additional sourcing context. Researchers comparing peptide purity standards should also consult resources on Bachem reference standards and peptide benchmarks when evaluating research-grade materials.
Conclusion
The Retatrutide Structural Mechanism: What Cryo-EM Reveals About Triple-Receptor Agonism comes down to a single engineered alpha-helix that speaks three receptor languages simultaneously. Cryo-EM has made it possible to see exactly how the peptide's N-terminal segment activates each receptor's transmembrane core while its C-terminal end navigates receptor-specific extracellular differences. Non-canonical amino acids provide enzymatic stability and receptor selectivity, while the fatty acid side chain extends circulating half-life to a clinically practical range.
For researchers working in this space, the actionable steps are clear: examine the structural data to understand why potency ratios were calibrated the way they were, compare retatrutide's binding architecture against earlier single and dual agonists, and track Phase 3 trial outcomes that will test whether structural advantages translate into durable clinical benefit. The cryo-EM data already provides a compelling molecular rationale for the efficacy signals observed so far.










Leave a Reply
Want to join the discussion?Feel free to contribute!