Retatrutide for Research: Mechanism, Structure, and GLP-1/GLP-3 Dual Action
A single investigational peptide producing near-bariatric levels of weight loss in a Phase 2 trial stopped the metabolic research community in its tracks. That peptide was retatrutide, and understanding Retatrutide for Research: Mechanism, Structure, and GLP-1/GLP-3 Dual Action has become one of the most urgent priorities in 2026 for scientists studying multi-receptor metabolic biology.
Key Takeaways
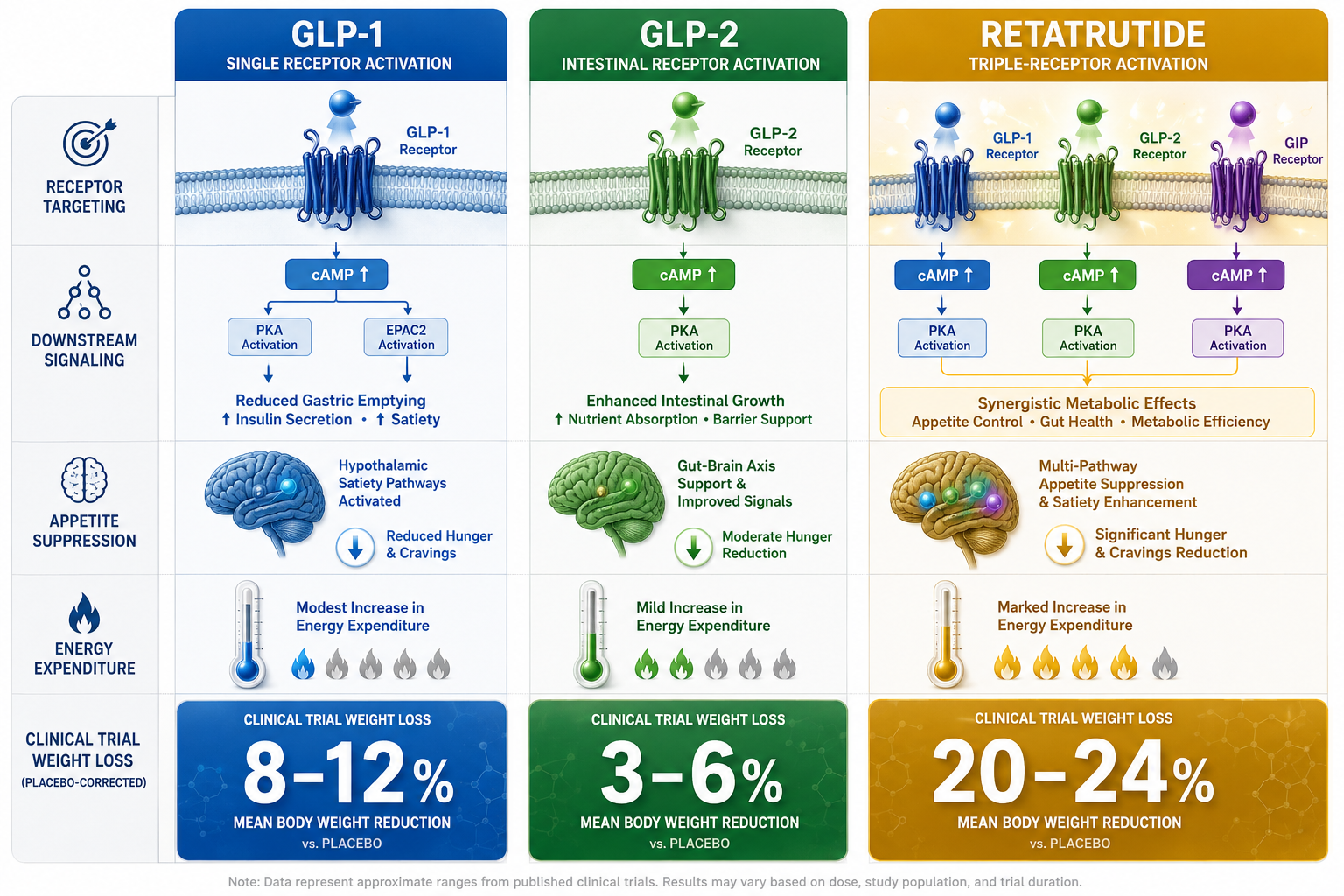
- Retatrutide is a triple receptor agonist targeting GLP-1R, GIPR, and GCGR simultaneously, not a simple dual GLP-1/GLP-3 agent.
- Its fatty-acid-modified structure enables a long half-life suitable for once-weekly dosing in research models.
- Receptor co-activation drives additive and potentially synergistic effects on energy balance, glucose regulation, and lipid metabolism.
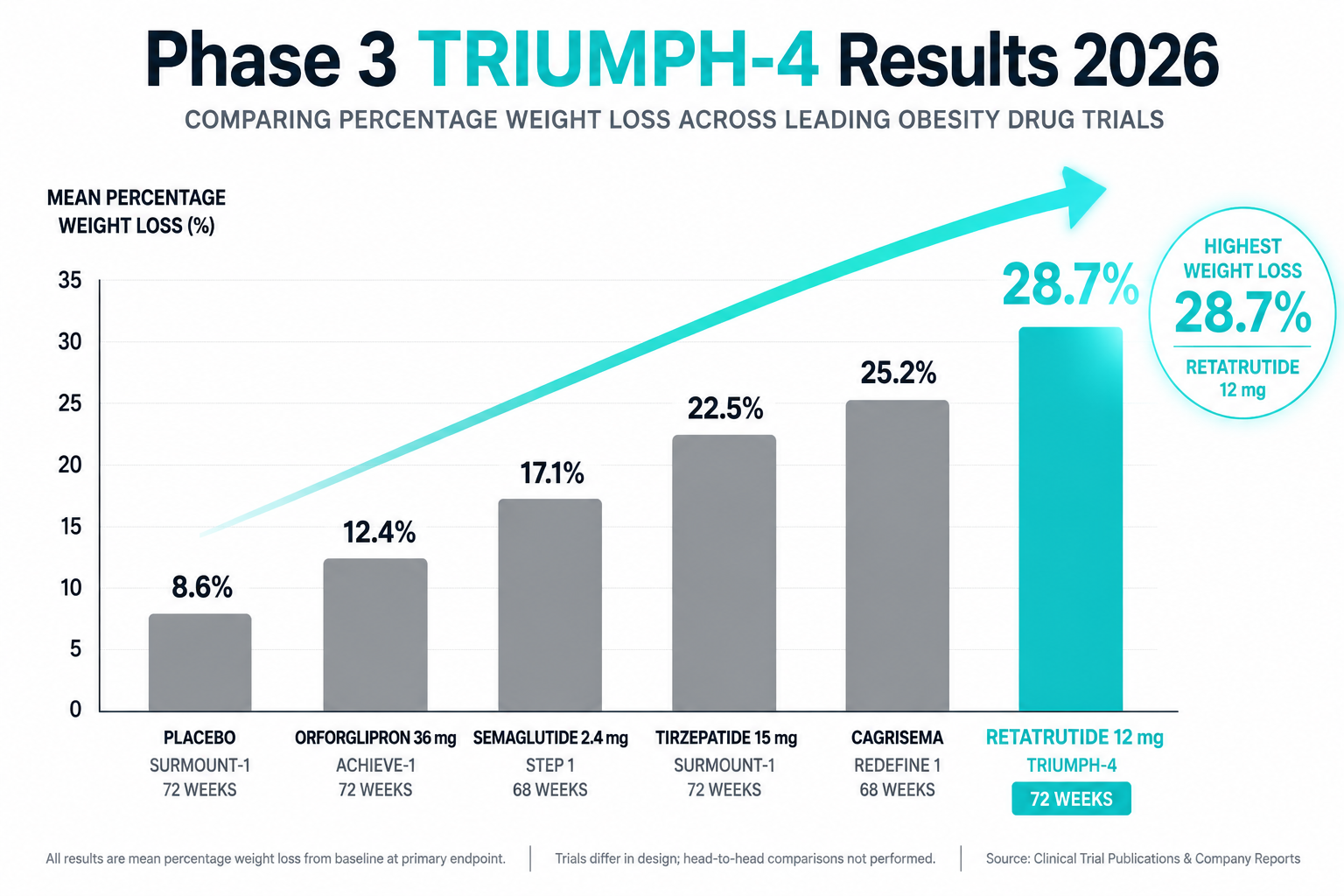
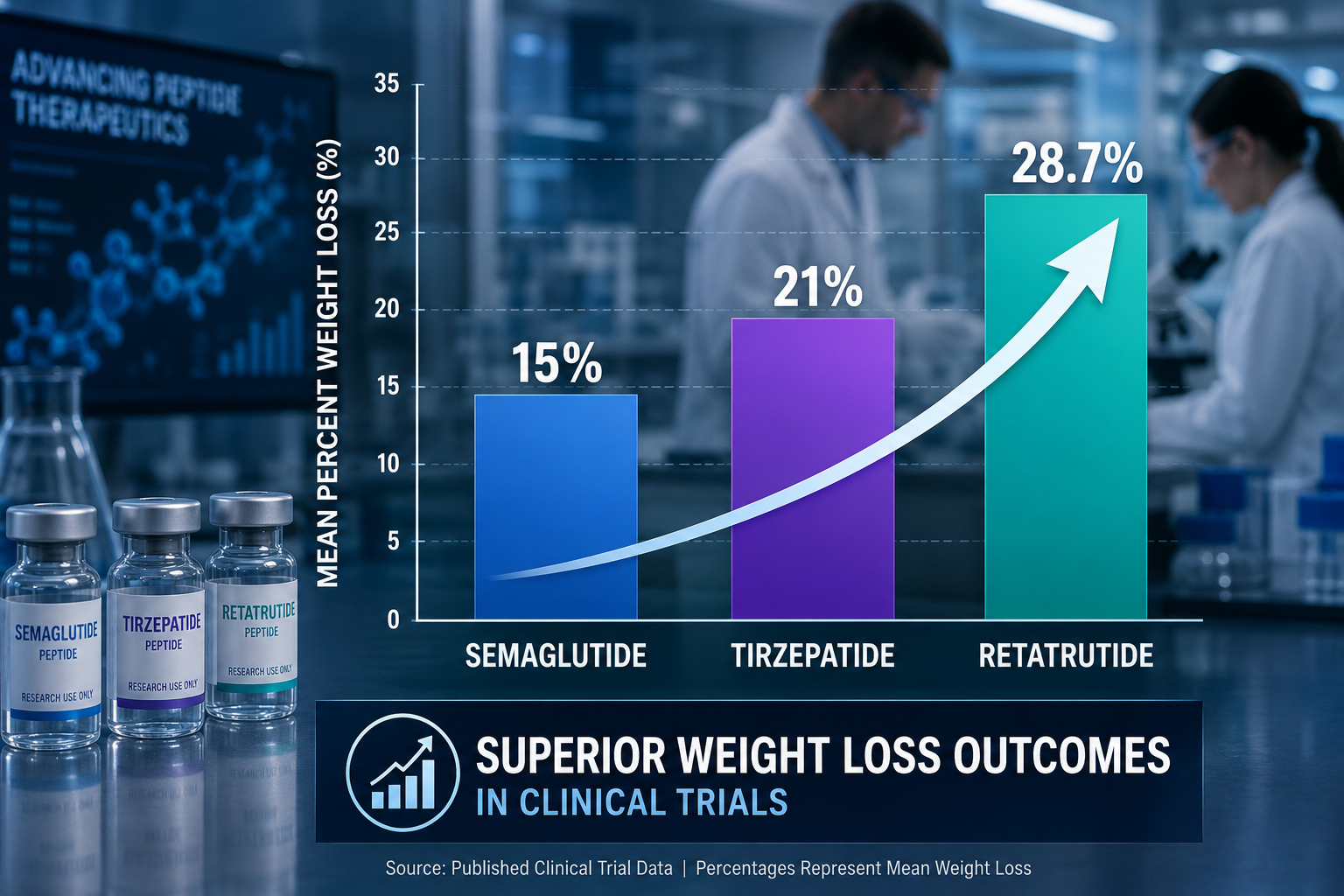
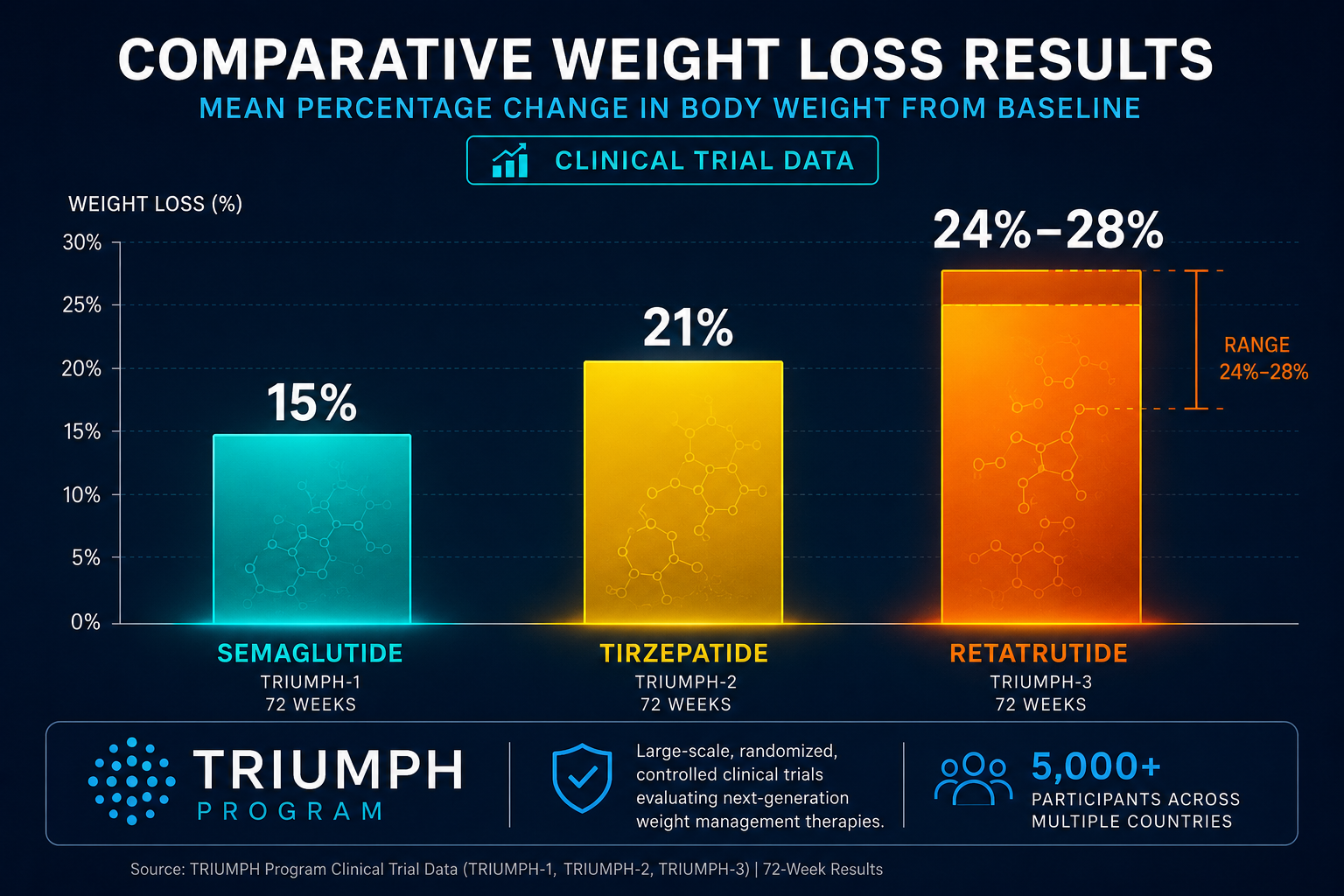
- Phase 2 data showed up to 24% body weight reduction; Phase 3 trials confirmed late-stage success in obesity and osteoarthritis pain endpoints in December 2025.
- Researchers tracking multi-agonist peptide science should understand both the structural basis and the downstream cAMP/PKA/EPAC signaling logic.
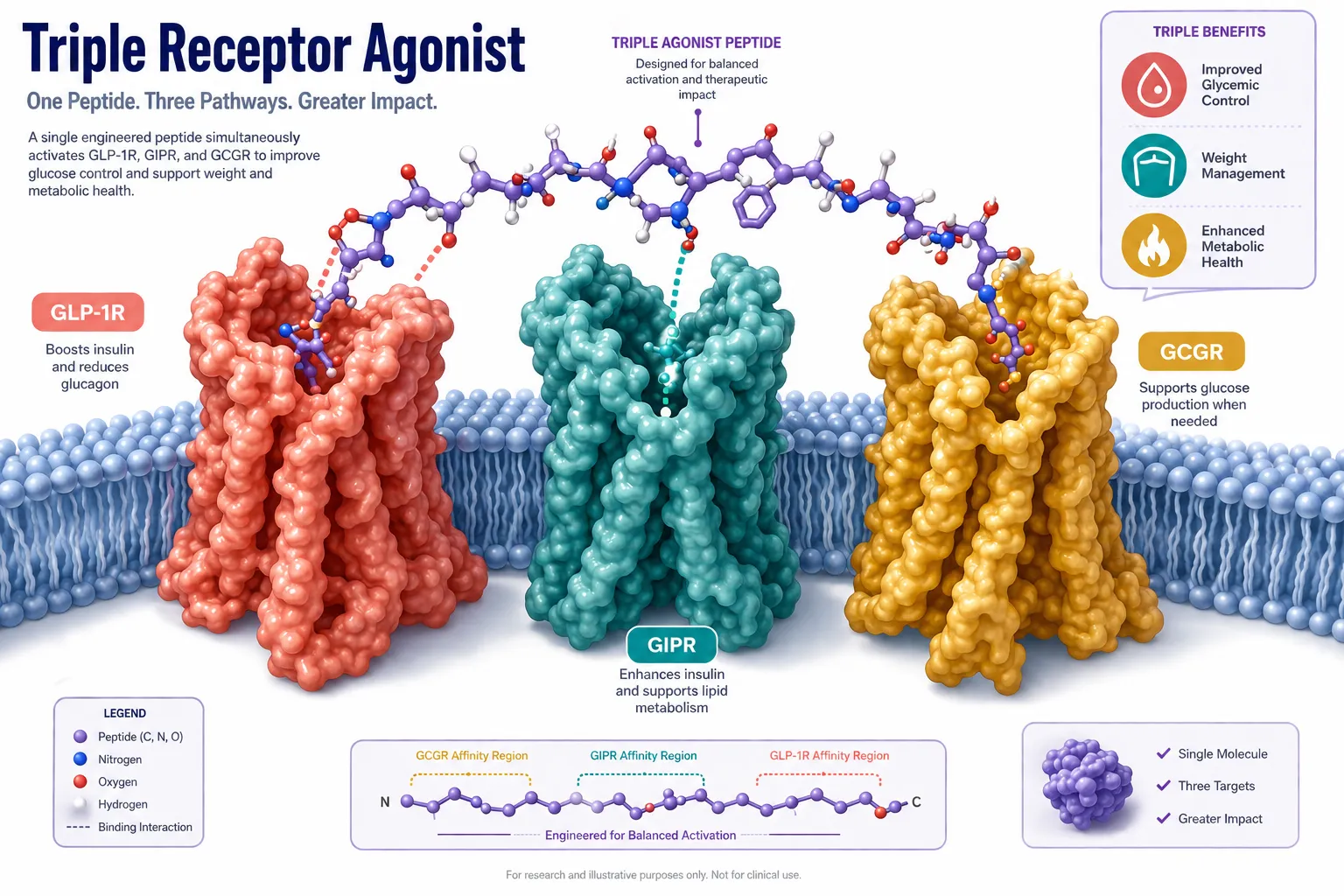
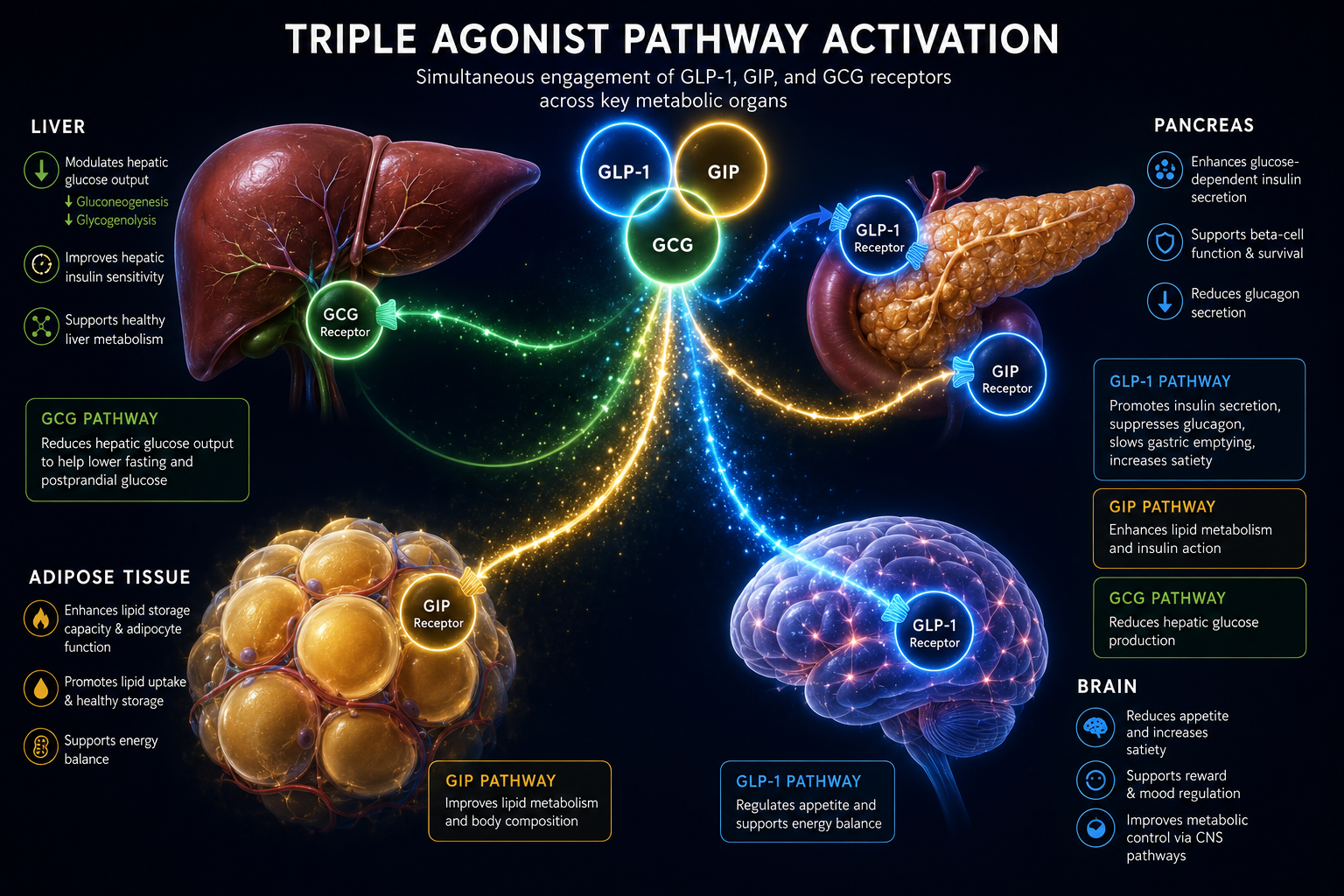
Molecular Structure: What Makes Retatrutide Unique
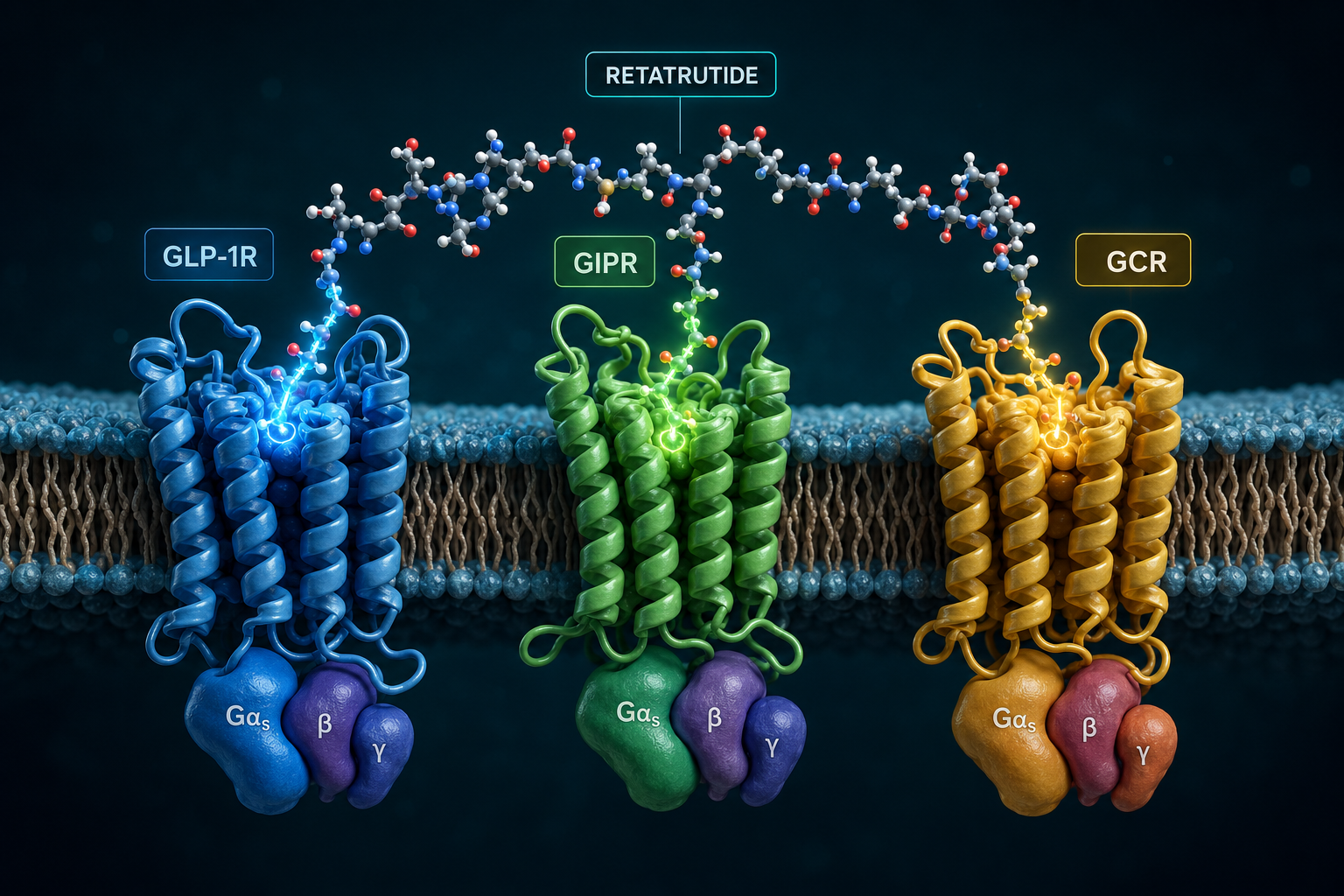
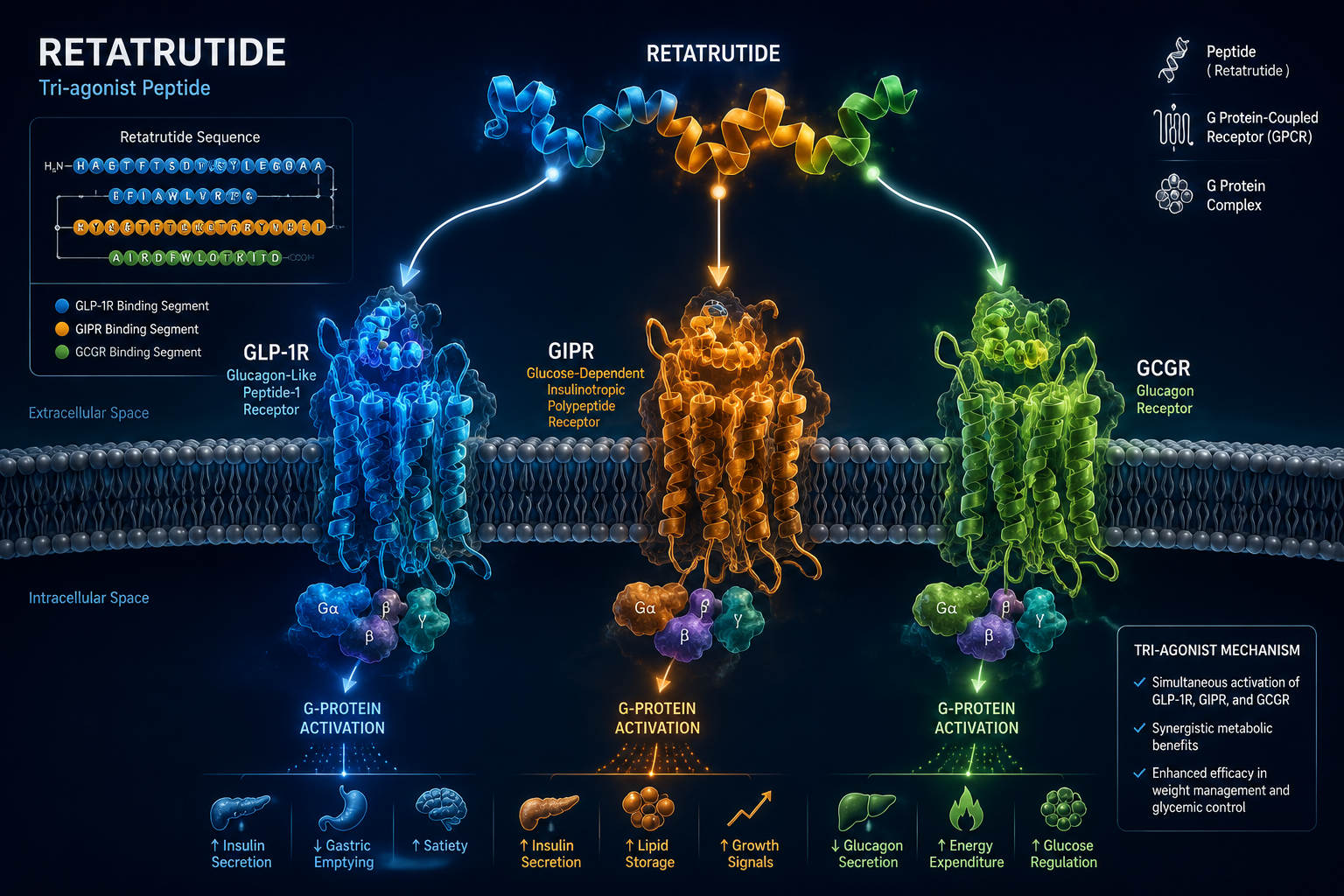
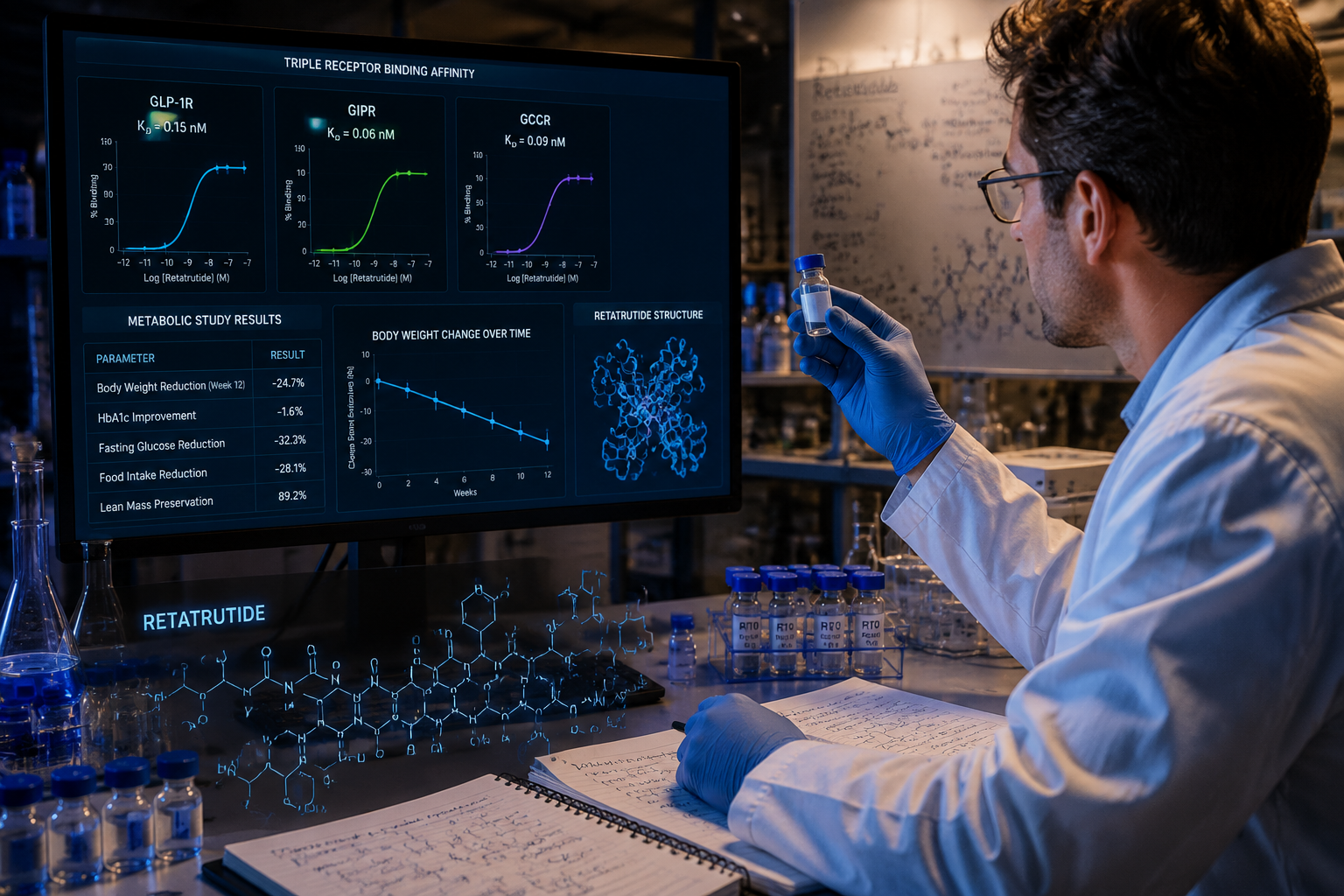
Retatrutide (LY3437943) is a 39-amino-acid synthetic peptide built on a modified glucagon backbone. Its design incorporates several deliberate structural features that set it apart from earlier incretin-based compounds.
Key structural elements include:
- A C18 fatty diacid chain attached via a linker to lysine at position 17, enabling albumin binding and extending plasma half-life to approximately 6 days.
- Strategic amino acid substitutions at positions 2 and 16 that confer resistance to dipeptidyl peptidase-4 (DPP-4) degradation.
- A C-terminal amide that stabilizes the peptide against exopeptidase activity.
- Balanced potency across all three target receptors rather than overwhelming selectivity for any single one.
This architecture is what allows researchers studying Retatrutide for Research: Mechanism, Structure, and GLP-1/GLP-3 Dual Action (and full triple agonism) to observe effects that neither a pure GLP-1 agonist nor a pure glucagon agonist could produce alone. For context on how earlier GLP-1 receptor agonists were structured, the GLP-1 incretin research overview provides useful background.
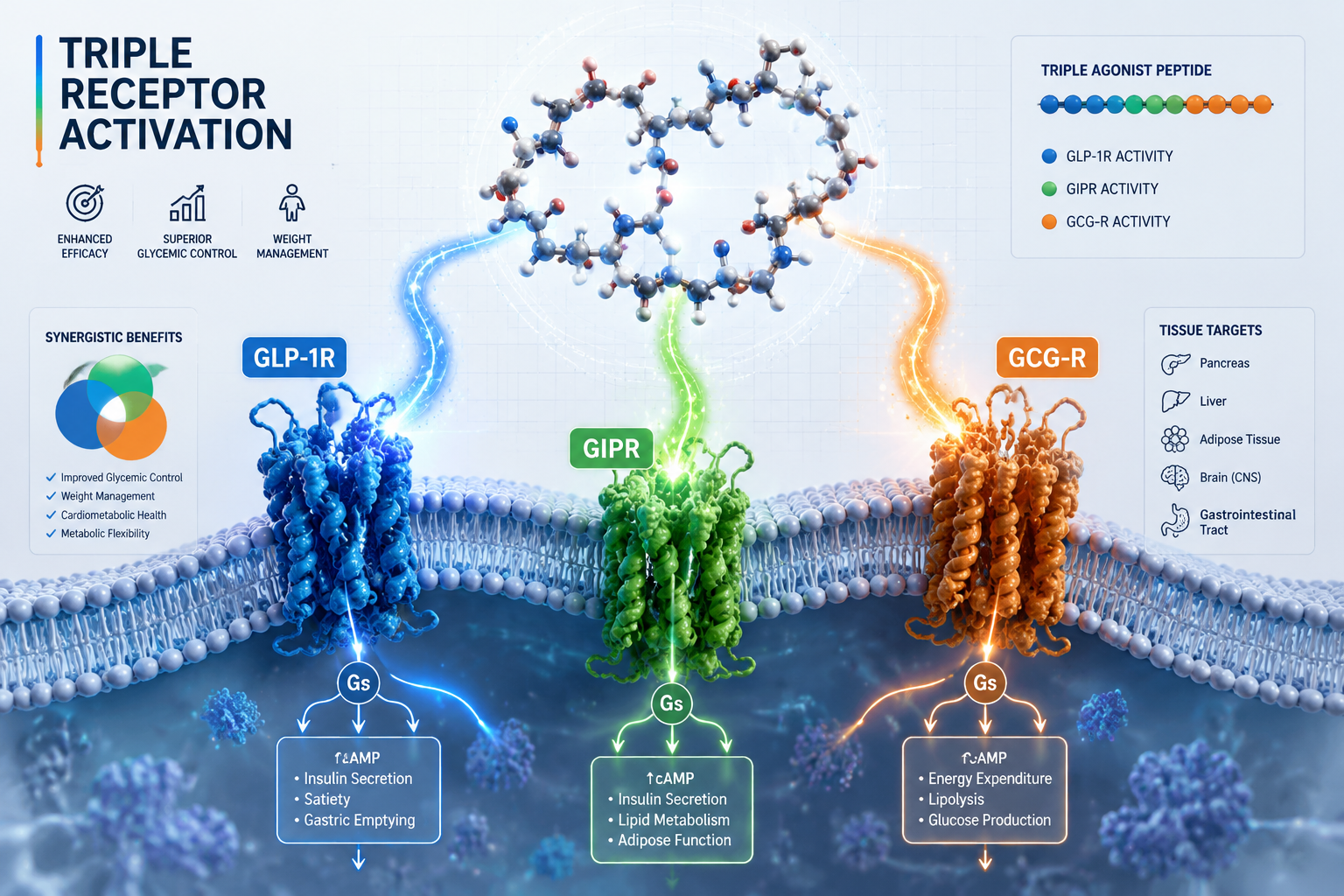
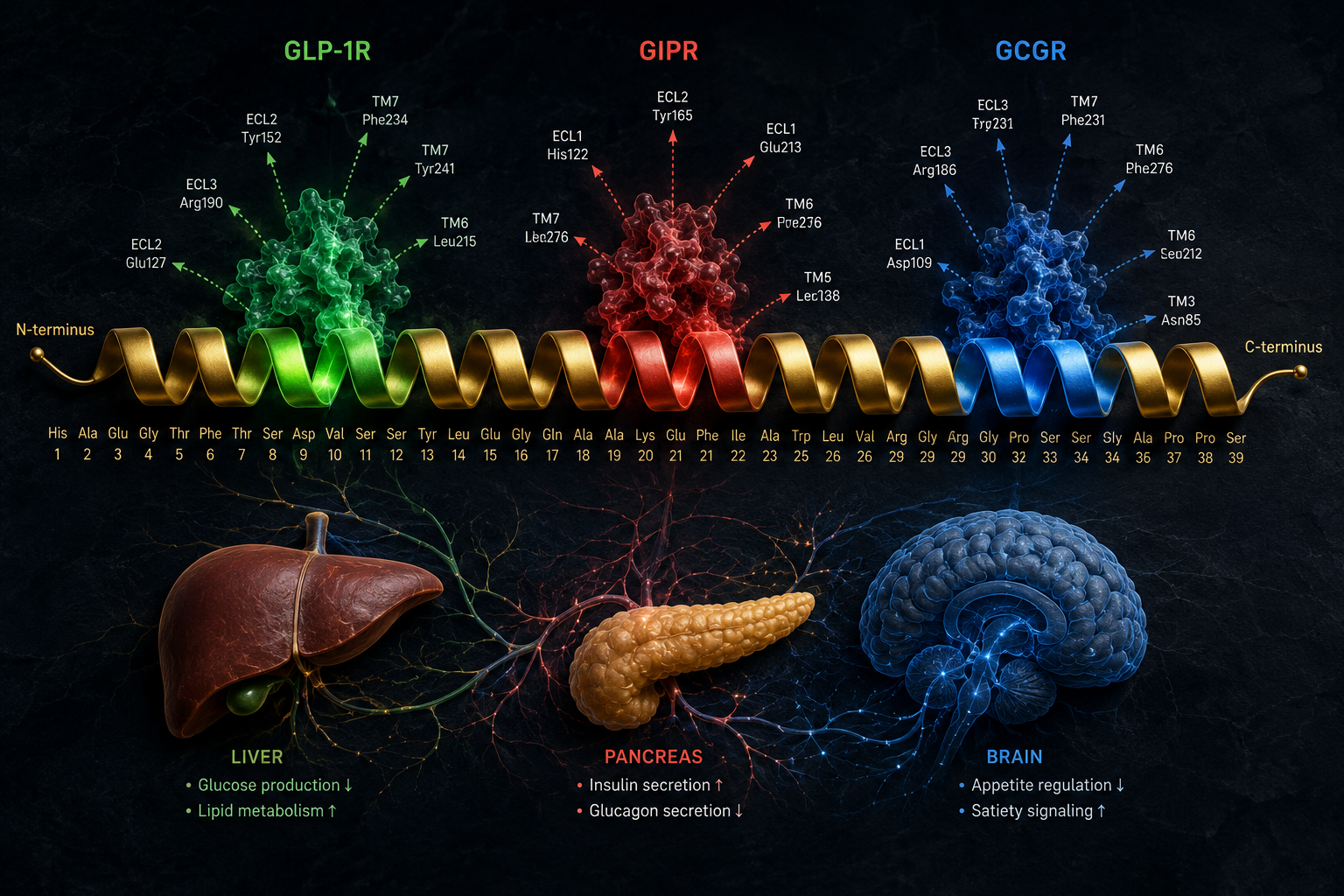
Receptor Potency Profile
| Receptor | Target | Primary Research Role |
|---|---|---|
| GLP-1R | Incretin axis | Insulin secretion, appetite suppression |
| GIPR | Glucose-dependent insulinotropic peptide | Insulin potentiation, fat cell signaling |
| GCGR | Glucagon receptor | Energy expenditure, hepatic lipid mobilization |
Cryo-EM structural studies have confirmed that retatrutide can engage all three receptor types, with the peptide adopting slightly different helical conformations depending on which receptor it occupies. This structural flexibility is central to its multi-target profile.
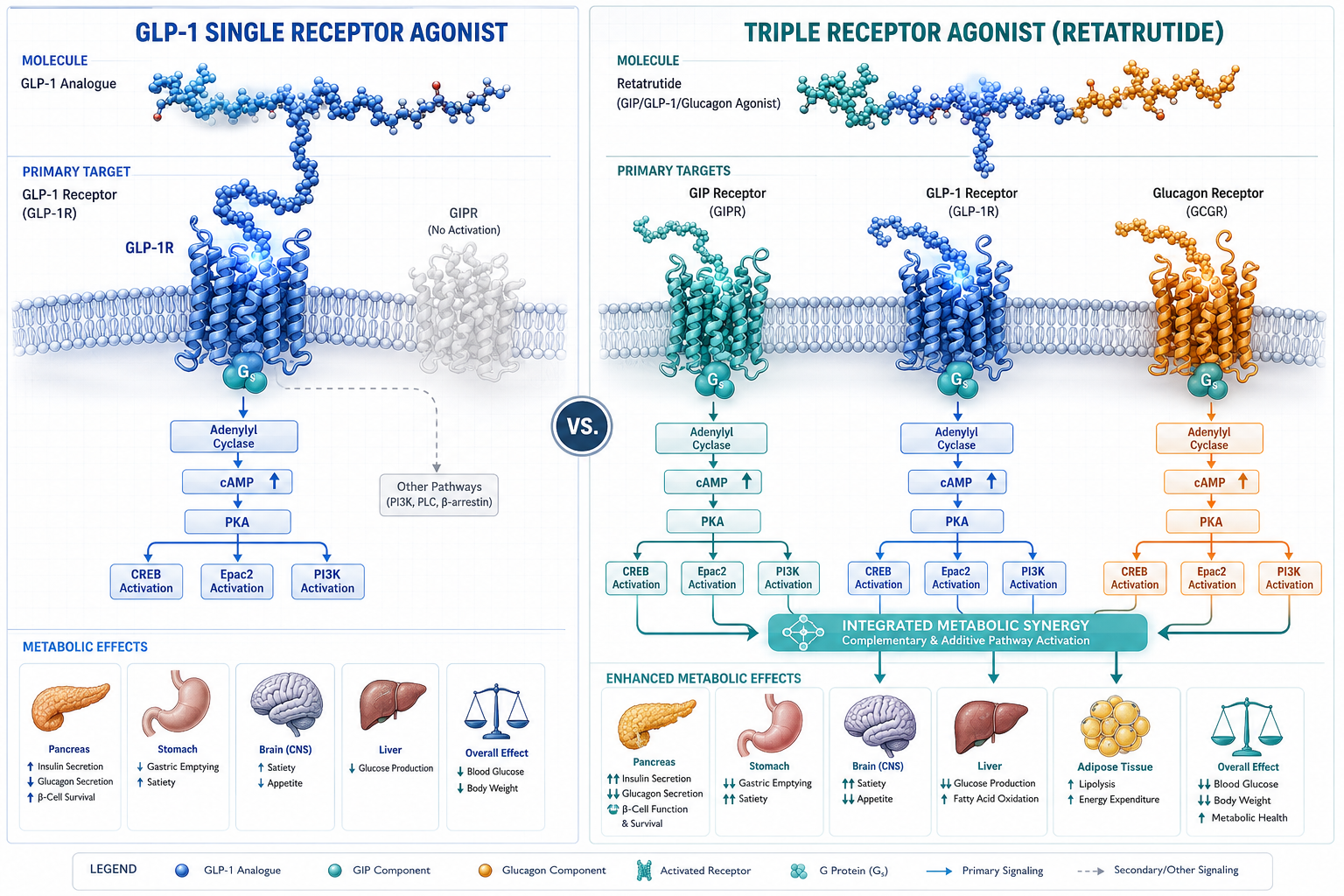
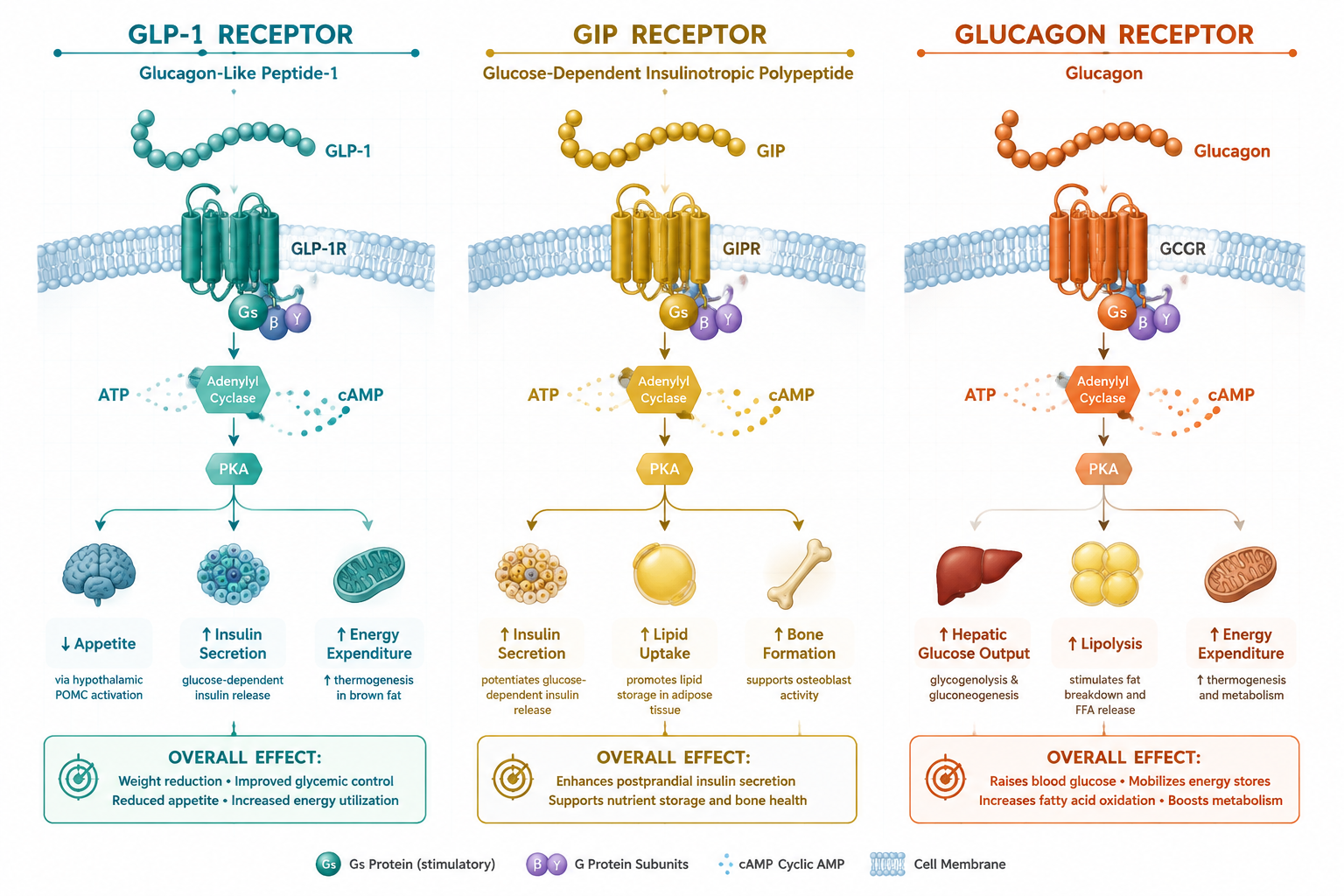
Cellular Signaling: cAMP, PKA, and EPAC Pathways
All three receptors targeted by retatrutide are G-protein-coupled receptors (GPCRs) that primarily signal through Gs proteins. When retatrutide binds, the shared downstream logic follows a defined cascade:
- Gs protein activation triggers adenylyl cyclase.
- Cyclic AMP (cAMP) accumulates intracellularly.
- cAMP activates two major effectors: protein kinase A (PKA) and exchange protein directly activated by cAMP (EPAC).
- PKA phosphorylates transcription factors and ion channels that regulate insulin gene expression and beta-cell survival.
- EPAC modulates vesicle exocytosis and cell adhesion signaling independently of PKA.
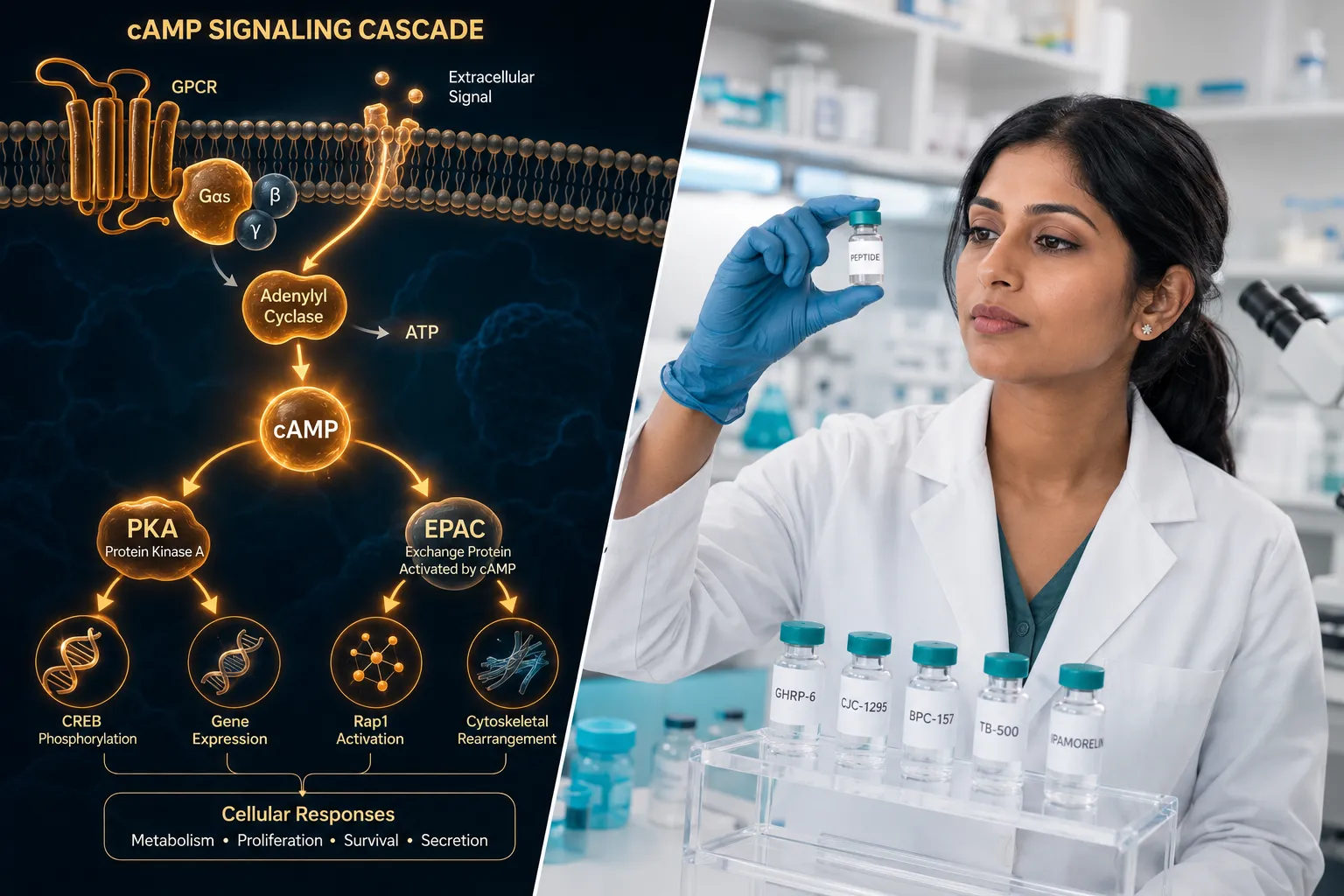
The simultaneous activation of GLP-1R, GIPR, and GCGR creates overlapping but non-identical cAMP pools in different tissue compartments. In pancreatic beta cells, GLP-1R and GIPR signals amplify insulin secretion. In adipose tissue, GIPR signaling modulates lipid storage. In the liver and brown adipose tissue, GCGR activation increases thermogenesis and fatty acid oxidation.
"The convergence of three receptor signals onto a shared cAMP axis, yet with tissue-specific outcomes, is what makes retatrutide a structurally elegant research tool for dissecting metabolic crosstalk."
This signaling architecture also explains why researchers interested in GLP-3 and retatrutide mechanisms find the compound particularly valuable: the interplay between incretin and glucagon arms of the pathway reveals metabolic biology that single-receptor tools cannot access.
For researchers also studying growth hormone secretagogues alongside metabolic peptides, the CJC-1295 with DAC research findings offer a complementary perspective on peptide half-life engineering.
Clinical Research Outcomes and Translational Significance
Understanding Retatrutide for Research: Mechanism, Structure, and GLP-1/GLP-3 Dual Action is inseparable from interpreting the clinical data that has validated the triple-agonist hypothesis.
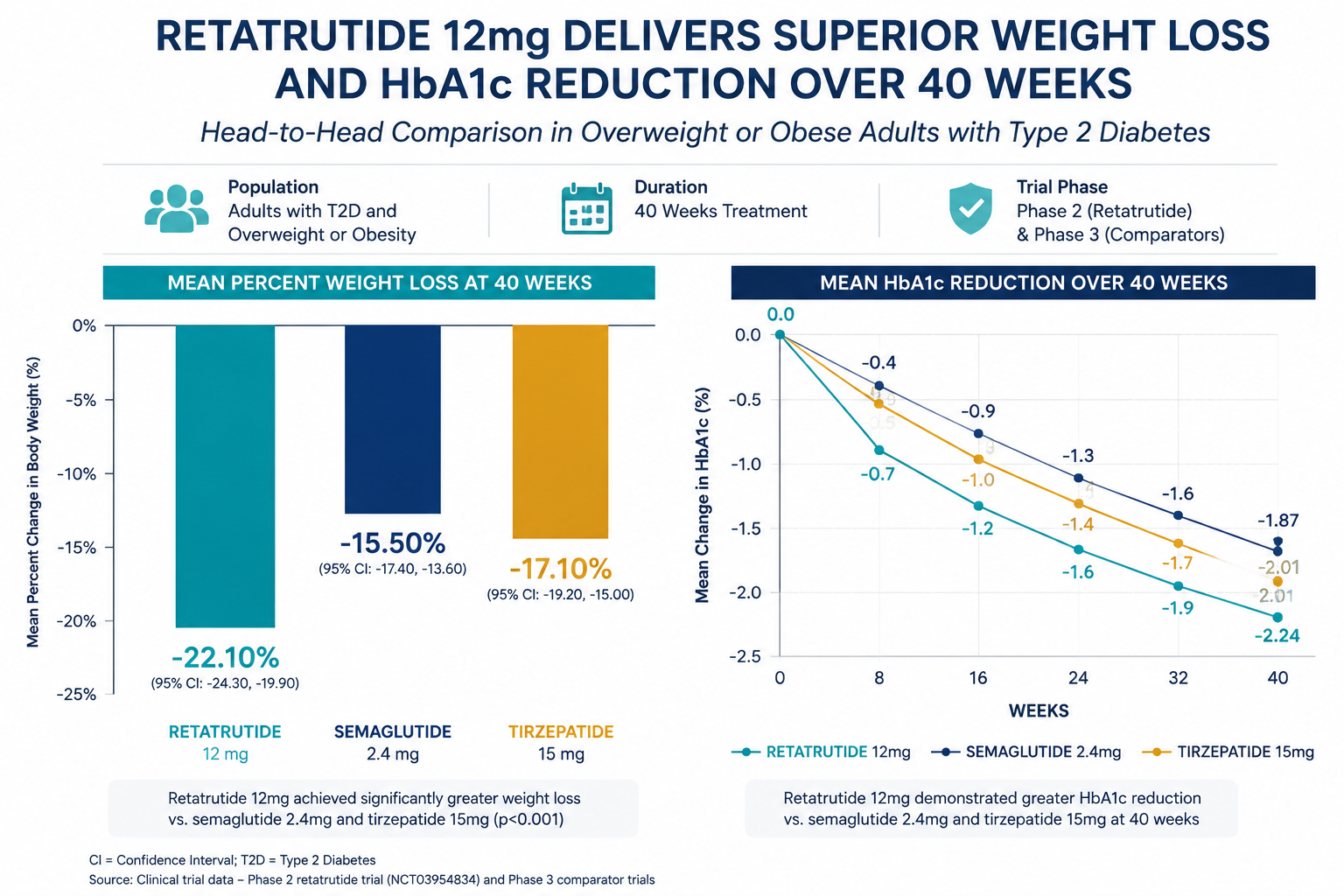
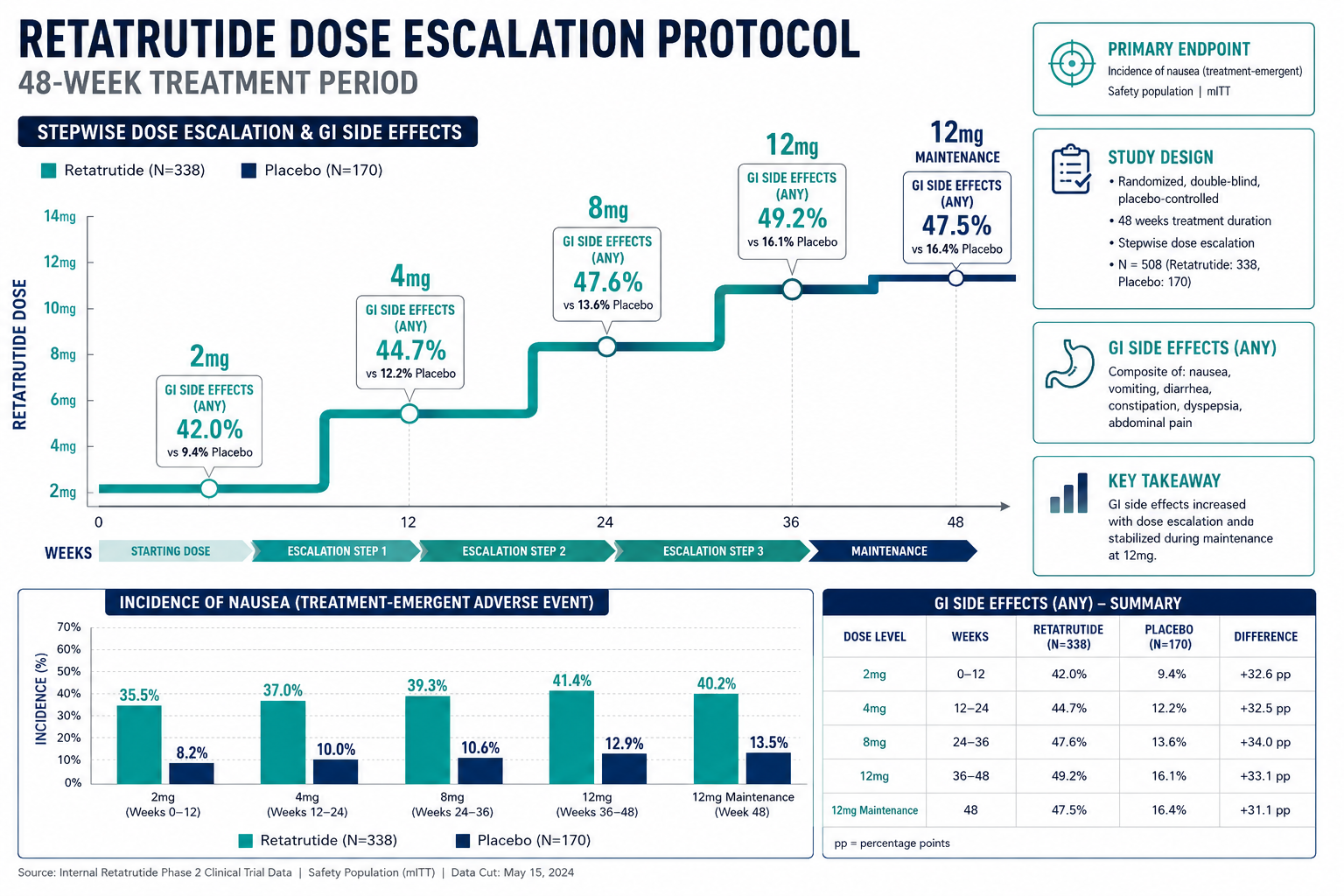
Phase 2 obesity trial (2023): Participants receiving the highest dose achieved approximately 24% mean body weight reduction over 48 weeks, a figure that approaches outcomes typically associated with bariatric surgery. This was substantially greater than what GLP-1 monotherapy had produced in comparable populations.
Phase 3 outcomes (December 2025): Late-stage trials confirmed statistically significant success across obesity endpoints and, notably, demonstrated meaningful reductions in osteoarthritis-related pain, an effect likely mediated through both weight-dependent joint offloading and direct anti-inflammatory receptor signaling.
Metabolic dysfunction-associated steatotic liver disease (MASLD): Preliminary data suggest retatrutide reduces hepatic fat fraction, consistent with the GCGR component driving hepatic lipid oxidation. This positions the compound as a research tool for liver biology as well as obesity science.

Researchers tracking the broader landscape of GLP-1 receptor agonist generations will recognize retatrutide as a structural and pharmacological leap beyond second-generation agents like semaglutide. Similarly, those following longevity peptide research may find the compound's metabolic and potentially cytoprotective signaling relevant to aging biology.
For researchers sourcing materials, the GLP-3 retatrutide 10mg research product is available for qualified laboratory use, and the Reta 10mg product tag provides additional sourcing information.
Conclusion
Retatrutide represents a structural and mechanistic milestone in peptide pharmacology. Its engineered triple-receptor profile, long half-life architecture, and convergent cAMP signaling logic make it one of the most information-rich research tools available for studying metabolic biology in 2026.
Actionable next steps for researchers:
- Review cryo-EM binding data to understand receptor-specific conformational differences before designing assay protocols.
- Map tissue-specific cAMP responses (beta cell vs. hepatocyte vs. adipocyte) to isolate receptor-arm contributions.
- Monitor ongoing Phase 3 data releases for MASLD and cardiovascular endpoints, which will clarify the full translational scope.
- Consider pairing retatrutide studies with complementary peptide tools, such as those covered in the cagrilintide and GLP-1 synergy research, to build multi-pathway metabolic models.
The structural nuances of retatrutide are not academic footnotes, they are the mechanistic foundation on which the next generation of metabolic therapeutics will be built.